

Struktur glikogen, sintesis, degradasi, fungsi
- 1691
- 299
- Julius Dibbert
Dia Glycogen Ia adalah karbohidrat penyimpanan kebanyakan mamalia. Karbohidrat biasanya dipanggil gula dan ini diklasifikasikan mengikut bilangan sisa yang disebabkan oleh hidrolisis (monosakarida, disakarida, oligosakarida dan polisakarida))).
Monosakarida adalah karbohidrat paling mudah yang diklasifikasikan mengikut bilangan karbon yang terkandung dalam struktur mereka. Terdapat triosas (3c), tetrosas (4c), pentosas (5c), hexosous (6c), heptosase (7c) dan octosas (8c).
Struktur glikogen kimia menunjukkan ikatan glikosid (sumber: glykogen.SVG: Kerja Derivatif Neurotoger: Marek M [Domain Awam] melalui Wikimedia Commons) Mengikut kehadiran kumpulan aldehid atau kumpulan cetona, monosakarida ini juga diklasifikasikan sebagai aldies atau ketosas masing -masing.
Disaccharides menimbulkan hidrolisis, dua monosakarida mudah, manakala oligosakarida menghasilkan 2 hingga 10 unit monosakarida dan polisakarida menghasilkan lebih daripada 10 monosakarida.
Glikogen adalah, dari sudut pandangan biokimia, polysaccharide yang terdiri daripada rantai bercabang dari aldose enam karbon, iaitu, heksosa yang dikenali sebagai glukosa. Secara grafik ia boleh diwakili kepada glikogen sebagai pokok glukosa. Ini juga dipanggil kanji haiwan.
Glukosa dalam tumbuh -tumbuhan disimpan sebagai kanji dan haiwan sebagai glikogen, yang disimpan terutamanya dalam tisu hati dan otot.
Di hati, glikogen dapat menubuhkan 10% jisimnya dan 1% jisim otot. Seperti pada lelaki 70kg, hati beratnya kira -kira 1800 g dan otot kira -kira 35 kg, jumlah glikogen otot jauh lebih besar daripada hepatik.
[TOC]
Struktur
Berat molekul glikogen boleh mencapai 108 g/mol, bersamaan dengan molekul glukosa 6 × 105. Glikogen terdiri daripada pelbagai rantai α-D-glikos bercabang. Glukosa (C6H12O6) adalah aldohexosa yang boleh diwakili dalam cara linear atau kitaran.
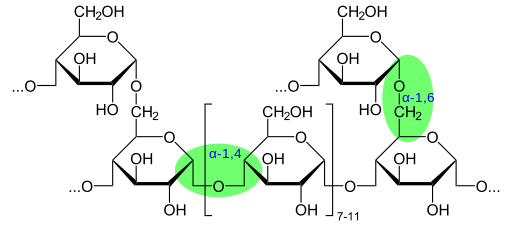
Glikogen mempunyai struktur yang sangat bercabang dan padat dengan rantai 12 hingga 14 sisa glukosa dalam bentuk α-D-glukosa yang dikaitkan dengan ikatan α- (1 → 4). Ramuan rantai dibentuk oleh pautan glukosid α- (1 → 6).
Glikogen, seperti kanji yang ditelan dalam diet, memberikan sebahagian besar karbohidrat yang diperlukan oleh tubuh. Dalam usus polysaccharides ini direndahkan oleh hidrolisis dan kemudian diserap ke arah torrent peredaran terutamanya sebagai glukosa.
Tiga enzim: ß-amylase, α-amylase dan amylo-α- (1 → 6) -glucosidase bertanggungjawab untuk degradasi usus kedua-dua glikogen dan kanji.
Α-amilase secara rawak menghidrolisiskan ikatan α-(1 → 4) rantai sampingan kedua-dua glikogen dan kanji, dan oleh itu menerima nama endoglysidase. Ss-amyla adalah exoglicosidase yang melepaskan ß-maltose dímeros memecahkan α- (1 → 4) pautan glikosid dari hujung rantai terluar tanpa mencapai ramuan.
Memandangkan fakta bahawa ß-amylase atau α-amylase merendahkan cawangan, produk akhir tindakannya adalah struktur yang sangat bercabang kira-kira 35 hingga 40 sisa glukosa yang dipanggil had dextrine.
Dextrin had akhirnya dihidrolisiskan pada titik cawangan yang mempunyai ikatan α- (1 → 6) melalui enzy-α- (1 → 6) -glucosidase, juga dikenali sebagai enzim "fitnah". Rantaian yang dikeluarkan oleh defloat ini selepas terdegradasi oleh ß-amilase dan α-amilase.
Oleh kerana glikogen yang ditelan memasuki glukosa, yang terdapat di dalam tisu mesti disintesis oleh organisma dari glukosa.
Boleh melayani anda: purin: ciri, struktur, fungsiSintesis
Sintesis glikogen dipanggil glikogenesis dan berlaku terutamanya dalam otot dan hati. Glukosa yang memasuki organisma dengan diet berlalu ke torrent peredaran darah dan dari sana di dalam sel, di mana ia segera difosforilasi oleh enzim yang dipanggil glycoquinase.
Fosforil glucoquinase hingga glukosa dalam karbon 6. ATP menyediakan fosforus dan tenaga untuk reaksi ini. Akibatnya, glukosa 6-fosfat dibentuk dan ADP dilepaskan. Kemudian, glukosa 6-fosfat menjadi glukosa 1-fosfat dengan tindakan fosfoglukomutase yang lumpur fosforus dari kedudukan 6 hingga kedudukan 1.
Glukosa 1-fosfat diaktifkan untuk sintesis glikogen, yang membayangkan penyertaan satu set tiga enzim lain: pyrophosphorylase UDP-glikose, sintetik glikogen dan amilo- (1.4 → 1.6) -Glicosyltransferase.
Glukosa-1-fosfat, bersama-sama dengan uridine trifosphat (UTP, nukleosida uridine triphosphate) dan dengan tindakan UDP-glycose-pyrophosphorylase, membentuk kompleks uridin diphosfat-glukosa (UDP GLC) (UDP GLC) (UDP GLC). Dalam proses ion pirofosfat dihidrolisis.
Kemudian, enzim glikogen sintetik membentuk ikatan glukosid antara C1 dari kompleks GLC UDP dan C4 dari residu terminal glukosa glikogen, dan UDP UDP dibebaskan dari kompleks glukosa diaktifkan UDP. Untuk tindak balas ini berlaku, mesti ada molekul glikogen yang sedia ada yang dipanggil "Glikogen Utama".
Glikogen primordial disintesis pada protein priming, glikogenin, yang mempunyai 37 kDa dan glysila dalam sisa tirosin menggunakan kompleks GLC UDP. Dari situ mereka dihubungkan sisa α-D-glukosa dengan 1 → 4 pautan dan rantai kecil terbentuk di mana tindakan glikogen synthesase.
Sebaik sahaja rantaian awal menghubungkan sekurang -kurangnya 11 residu glukosa, enzim cawangan atau amile- (1.4 → 1,6) -glicosyltransferase memindahkan sekeping rantai 6 atau 7 sisa glukosa ke rantai bersebelahan dalam kedudukan 1 → 6, yang menetapkan cawangan titik. Molekul glikogen yang dibina itu berkembang dengan penambahan unit glukosa dengan pautan glikosid 1 → 4 dan lebih banyak kesan.
Degradasi
Degradasi glikogen dipanggil glucogenolysis, dan tidak setara dengan laluan terbalik sintesisnya. Kelajuan laluan ini dibatasi oleh kelajuan tindak balas yang dipangkin oleh glikogen phosphorylase.
Fosfora glikogen bertanggungjawab untuk pemisahan (fosforolisis) pautan 1 → 4 dari rantai glikogen, melepaskan glukosa 1-fosfat. Tindakan enzimatik bermula di hujung rantai paling luar dan dikeluarkan secara berurutan sehingga 4 sisa glukosa kekal di setiap sisi dari kesan.
Kemudian, enzim lain, α- (1 → 4) → α- (1 → 4) glucano transferas, meninggalkan titik cawangan yang terdedah dengan memindahkan unit trisakarida dari satu cawangan ke cawangan lain. Ini membolehkan amilo- (1 → 6) -glucosidase (enzim unrampic). Tindakan gabungan enzim ini berakhir sepenuhnya berpecah kepada glikogen.
Oleh kerana reaksi awal fosfomutase boleh diterbalikkan, glukosa 6-fosfat boleh dibentuk dari residu glukosa 1-fosfat dibahagi dari glikogen. Di dalam hati dan buah pinggang, tetapi tidak di dalam otot, terdapat enzim, glukosa-6-phosphatase, mampu berkumpul hingga glukosa 6-fosfat dan mengubahnya menjadi glukosa percuma.
Boleh melayani anda: fotolisisGlukosa defosforilasi dapat merebak ke darah, dan itulah bagaimana glikogenolisis hepatik dicerminkan dalam peningkatan nilai glukosa darah (glikemia).
Peraturan sintesis dan degradasi
Sintesis
Proses ini dijalankan pada dua enzim asas: synthesase glikogen dan fosforilase glikogen, sehingga apabila salah satu daripadanya diaktifkan, yang lain berada dalam keadaan tidak aktifnya. Peraturan ini menghalang tindak balas menentang sintesis dan kemerosotan dari berlaku secara serentak.
Bentuk aktif dan bentuk enzim yang tidak aktif sangat berbeza, dan interversi bentuk fosforilase aktif dan tidak aktif dan glikogen sintetik tertakluk kepada kawalan hormon yang ketat.
Adrenalin adalah hormon yang dilepaskan dari sumsum adrenal, dan glukagon adalah satu lagi yang berlaku di bahagian endokrin dari pankreas. Pankreas endokrin menghasilkan insulin dan glukagon. Langerhans Islet α adalah yang mensintesis glukagon.
Adrenalin dan glukagon adalah dua hormon yang dibebaskan apabila tenaga diperlukan sebagai tindak balas kepada penurunan tahap glukosa darah. Hormon ini merangsang pengaktifan fosforilas glikogen dan menghalang glikogen synthesase, dengan itu merangsang glikogenolisis dan menghalang glikogenesis.
Walaupun adrenalin melakukan tindakannya terhadap otot dan hati, glukagon hanya bertindak pada hati. Hormon ini disertai dengan reseptor membran tertentu di sel putih, yang mengaktifkan adenilasi siklasa.
Pengaktifan adenilat siklase memulakan air terjun enzimatik yang, di satu pihak, mengaktifkan proteinquinase yang bergantung kepada AMPC yang tidak aktif kepada glikogen sintetik dan mengaktifkan fosforilasi glikogen oleh fosforilasi (secara langsung dan tidak langsung).
Otot rangka mempunyai mekanisme lain untuk mengaktifkan fosforilas glikogen melalui kalsium, yang dilepaskan sebagai akibat dari depolarisasi membran otot pada permulaan penguncupan.
Degradasi
Air terjun enzimatik yang diterangkan di atas akhirnya meningkatkan tahap glukosa dan apabila mereka mencapai tahap tertentu, glikogenesis diaktifkan dan glucogenolysis dihalang, juga menghalang pembebasan adrenalin dan glukagon selanjutnya.
Glikogenesis diaktifkan dengan pengaktifan phosphorylase phosphatase, enzim yang mengawal sintesis glikogen oleh beberapa mekanisme, yang menyiratkan inaktivasi fosforilase kinase dan phosphorylase α, yang merupakan inhibitor sintosise glikogen.
Insulin menggalakkan kemasukan glukosa ke dalam sel-sel otot, meningkatkan tahap glukosa 6-fosfat, yang merangsang defosforilasi dan pengaktifan sintesis glikogen. Oleh itu sintesis bermula dan degradasi glikogen dihalang.
Fungsi
Glikogen otot membentuk rizab tenaga untuk otot yang, seperti lemak rizab, membolehkan otot untuk memenuhi fungsinya. Menjadi sumber glukosa, glikogen otot digunakan semasa bersenam. Tempahan ini meningkat dengan latihan fizikal.
Di hati, glikogen juga merupakan sumber rizab penting untuk kedua -dua fungsi organ dan untuk sumbangan glukosa ke seluruh badan.
Fungsi glikogen hati ini disebabkan oleh fakta bahawa hati mengandungi glukosa 6-phosphatase, enzim yang mampu menghapuskan kumpulan fosfat glukosa 6-fosfat dan mengubahnya menjadi glukosa bebas. Glukosa percuma, tidak seperti glukosa fosforilasi, boleh disebarkan melalui membran hepatosit (sel hati).
Ia boleh melayani anda: sporulasi: dalam tumbuhan, dalam kulat dan bakteriaIni adalah bagaimana hati dapat memberikan glukosa kepada peredaran dan mengekalkan tahap glukosa yang stabil, walaupun dalam keadaan puasa yang berpanjangan.
Fungsi ini sangat penting, kerana otak dipelihara hampir secara eksklusif dari glukosa darah, hipoglikemia yang teruk (kepekatan glukosa darah yang sangat rendah) dapat menyebabkan kehilangan pengetahuan.
Penyakit berkaitan
Penyakit berkaitan glikogen menerima nama generik "Penyakit Penyimpanan Glikogen".
Penyakit ini merupakan sekumpulan patologi keturunan yang dicirikan oleh deposit dalam tisu -tisu yang tidak normal atau jenis glikogen.
Sebilangan besar penyakit penyimpanan glikogen disebabkan oleh defisit sifat genetik mana -mana enzim yang terlibat dalam metabolisme glikogen.
Mereka diklasifikasikan kepada lapan jenis, yang kebanyakannya mempunyai nama mereka sendiri dan masing -masing dihasilkan oleh defisit enzimatik yang berbeza. Ada yang fana dalam peringkat awal kehidupan, sementara yang lain disertai dengan kelemahan otot dan defisit semasa bersenam.
Contoh yang luar biasa
Beberapa penyakit berkaitan glikogen yang paling menonjol adalah seperti berikut:
- Penyakit Von Gierke atau Penyakit Penyimpanan Glikogen Jenis I, dihasilkan oleh defisit glukosa 6-fosfatase di hati dan buah pinggang.
Ia dicirikan oleh pertumbuhan hati yang tidak normal (hepatomegali) kerana pengumpulan glikogen dan hipoglikemia yang dibesar -besarkan, kerana hati tidak dapat memberikan glukosa kepada peredaran. Pesakit dengan keadaan ini mempunyai perubahan pertumbuhan.
- Penyakit Pompe atau Jenis II disebabkan oleh α-defisit (1 → 4) -glucano 6-glikosiltransferas di otot hati, jantung dan rangka. Penyakit ini, seperti Andersen atau Jenis IV, mematikan sebelum dua tahun hidup.
- Penyakit McArdle atau jenis V mempunyai defisit fosforilas otot dan disertai dengan kelemahan otot, penurunan toleransi senaman, pengumpulan abnormal glikogen otot dan ketiadaan laktat semasa bersenam semasa bersenam.
Rujukan
- Bhattacharya, k. (2015). Penyiasatan dan Pengurusan Penyakit Penyimpanan Glikogen Hepatik. Pediatrik translasi, 4(3), 240-248.
- Dagli, a., Sentner, c., & Weinstein, D. (2016). Jenis Penyakit Penyimpanan Glikogen III. Ulasan Gene, 1-16.
- Guyton, a., & Hall, J. (2006). Buku teks fisiologi perubatan (Edisi ke -11.). Elsevier Inc.
- Mathews, c., Van holde, k., & Ahern, k. (2000). Biokimia (Edisi ke -3.). San Francisco, California: Pearson.
- McKiernan, ms. (2017). Pathobiologi keinginan penyimpanan glikogen hepatik. Curr Pathobiol Rep.
- Murray, r., Bender, d., Botham, k., Kennelly, ms., Rodwell, v., & Weil, p. (2009). Biokimia Illustrated Harper (Edisi ke -28.). McGraw-Hill Medical.
- Nelson, d. L., & Cox, m. M. (2009). Prinsip Biokimia Lehninger. Edisi Omega (Edisi ke -5.).
- Rawn, j. D. (1998). Biokimia. Burlington, Massachusetts: Penerbit Neil Patterson.
- Tarnopolsky, m. Ke. (2018). Myopathies yang berkaitan dengan gangguan metabolisme glikogen. Neurotherapeutics.
- « Sejarah Argon, Struktur, Hartanah, Kegunaan
- Fungsi Bijektif Apa itu, Bagaimana Ia Dilakukan, Contoh, Latihan »