

Ciri -ciri liang nuklear, fungsi, komponen
- 4236
- 179
- Erick Krajcik
Dia liang nuklear (Dari bahasa Yunani, liang = langkah atau transit) adalah "pintu" nukleus yang membolehkan kebanyakan pengangkutan antara nukleoplasma dan sitoplasma. Liang nuklear menyertai membran dalaman dan luaran nukleus untuk membentuk saluran, yang berfungsi untuk pengangkutan protein dan RNA.
Perkataan poro tidak mencerminkan kerumitan struktur yang dipersoalkan. Kerana ini, lebih baik merujuk kepada kompleks liang nuklear (CPN), bukannya liang nuklear. CPN mungkin mengalami perubahan dalam strukturnya semasa pengangkutan atau keadaan kitaran sel.
Baru -baru ini, telah ditemui bahawa nukleoporin, protein yang membentuk CPN, mempunyai peranan penting dalam mengawal ekspresi gen. Oleh itu, apabila mutasi berlaku yang mempengaruhi fungsi nukleoporin, patologi dihasilkan pada manusia, seperti autoimun, kardiomiopati, jangkitan virus dan kanser.
[TOC]
Ciri -ciri
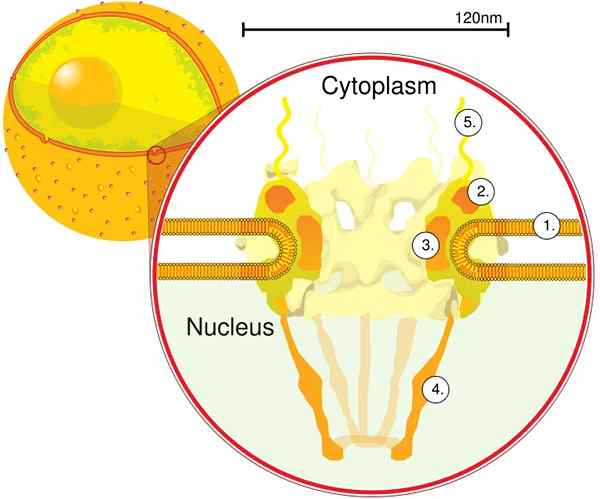
Melalui tomografi elektron, ditentukan bahawa CPN mempunyai ketebalan 50 nm, diameter luaran antara 80 dan 120 nm, dan diameter dalaman 40 nm. Molekul besar seperti subunit besar ribosom (PM 1590 kDa) boleh dieksport di luar nukleus melalui CPN. Dianggarkan terdapat antara tahun 2000 dan 4000 cpn setiap nukleus.
Berat molekul CPN individu adalah kira -kira antara 120 dan 125 mda (1 mda = 106 Da) dalam vertebrata. Sebaliknya, CPN lebih kecil dalam ragi, di mana ia mempunyai kira -kira 60 mda. Walaupun saiz besar CPN, nukleoporin sangat dipelihara dalam semua eukariota.
Translocalization melalui CPN adalah proses yang cepat, yang kelajuannya adalah 1000 translocations/saat. Walau bagaimanapun, CPN tidak menentukan arah aliran pengangkutan itu sendiri.
Ini bergantung kepada kecerunan RangTP, yang lebih besar dalam nukleus daripada di sitosol. Kecerunan ini dikekalkan oleh faktor penukar guanine berlari.
Semasa kitaran sel, CPN pengalaman pemasangan dan kitaran perpecahan. Perhimpunan berlaku di antara muka dan sejurus selepas mitosis.
Fungsi
Asid Ribonucleic (RNA nuklear kecil, RNA messenger, RNA pemindahan), protein dan ribonukleoprotein (RNP) mesti diangkut secara aktif melalui CPN. Ini bermakna tenaga hidrolisis ATP dan GTP diperlukan. Setiap molekul diangkut dengan cara tertentu.
Secara umum, molekul RNA dibungkus dengan protein yang membentuk kompleks RNP, dieksport dengan cara ini. Sebaliknya, protein yang secara aktif diangkut ke nukleus mesti mempunyai tanda urutan lokasi dalam nukleus (SLN), pemilik sisa asid amino dengan beban positif (contohnya, kKKRK).
Boleh melayani anda: mesosomProtein yang dieksport ke nukleus mesti mempunyai isyarat eksport ke nukleus (NES) yang kaya dengan asid amino leucine.
Di samping memudahkan pengangkutan antara nukleus dan sitoplasma, CPN terlibat dalam organisasi kromatin, peraturan ekspresi gen dan pembaikan DNA. Nukleoporin (NUPS) Menggalakkan pengaktifan atau penindasan transkripsi, tanpa mengira keadaan percambahan sel.
Dalam ragi, nupts terdapat di CNP pembungkus nuklear. Di metazoaries mereka berada di dalam. Mereka menjalankan fungsi yang sama dalam semua eukariot.
Import bahan
Melalui CPN terdapat penyebaran pasif molekul kecil di kedua -dua arah dan pengangkutan aktif, import protein, eksport RNA dan ribonukleoprotein (RNPs), dan pesawat molekul dua arah. Yang terakhir termasuk RNA, RNP dan protein yang terlibat dalam isyarat, biogenesis dan penggantian.
Pengimportan protein dalam nukleus berlaku dalam dua langkah: 1) protein yang mengikat ke sisi sitoplasma CPN; 2) Translocalization bergantung ATP melalui CPN. Proses ini memerlukan hidrolisis ATP dan pertukaran GTP/KDNK antara nukleus dan sitoplasma.
Menurut model pengangkutan, kompleks protein penerimaan bergerak di sepanjang saluran melalui kesatuan, pemisahan dan kesatuan lagi ke urutan FG berulang nukleoporin. Dengan cara ini, kompleks bergerak dari satu nukleoporin ke yang lain di dalam CPN.
Eksport bahan
Sama dengan pengimportan. Berlari GTPase mengenakan arah ke arah pengangkutan melalui CNP. RAN adalah suis molekul dengan dua keadaan konformasi, bergantung kepada sama ada ia dikaitkan dengan KDNK atau GTP.
Dua protein pengawalseliaan tertentu mencetuskan penukaran antara kedua-dua negeri: 1) Cytosolic GTPase mengaktifkan protein (GAP), yang menghasilkan hidrolisis GTP dan dengan itu menukarkan RAN-GTP ke RAN-KDNK; dan 2) Faktor Pertukaran Guanina Nuklear (GEF), yang menggalakkan pertukaran KDNK oleh GTP dan menukarkan RAN-KDNK ke RAN-GTP.
Cytosol terutamanya mengandungi RAN-KDNK. Nukleus terutamanya mengandungi RAN-GTP. Kecerunan ini dari dua bentuk konformasi RAN mengarahkan pengangkutan di alamat yang sesuai.
Pengimportan penerima, bersama-sama dengan kedudukan, difasilitasi oleh kesatuan kepada pengulangan-fg. Sekiranya anda sampai ke bahagian nuklear CNP, RAN-GTP menyertai penerima untuk melepaskan kedudukan anda. Oleh itu, RAN-GTP mewujudkan arah proses import.
Eksport nuklear adalah serupa. Walau bagaimanapun, Ran-GTP dalam nukleus menggalakkan kesatuan kedudukan ke penerima eksport. Apabila penerima eksport bergerak melalui liang ke arah sitosol, ia memenuhi jurang RAN, yang mendorong hidrolisis GTP ke KDNK. Akhirnya, penerima dibebaskan dari kedudukannya dan Ran-KDNK di sitosol.
Boleh melayani anda: leukosit polimorfonuklearPengangkutan RNA
Eksport beberapa jenis RNA adalah serupa dengan eksport protein. Contohnya, ARNT dan RNANS (Nuklear Kecil) Gunakan kecerunan RANGTP dan diangkut melalui CPN dengan cara. Eksport ribosom matang juga bergantung kepada kecerunan rangtp.
MRNA dieksport dengan cara yang sangat berbeza dari protein dan RNA lain. Untuk eksportnya, mRNA membentuk kompleks RNP Messenger (RNPM), di mana molekul RNM dikelilingi oleh beratus -ratus molekul protein. Protein ini mempunyai pemprosesan, Capping, Empalme dan Polyadentilly MNA.
Sel mesti dapat membezakan antara RNPM dengan RNM dan RNPM matang dengan mRNA yang tidak matang. MRNA, yang membentuk kompleks rpnm, boleh mengamalkan topologi yang perlu diubahsuai untuk pengangkutan. Sebelum RNPM memasuki CPN, langkah kawalan yang dijalankan oleh kompleks protein tramp dan exosome berlaku.
Apabila RNPM matang dipasang, RPNM diangkut melalui saluran dengan cara penerima pengangkutan (NXF1-NXT1). Penerima ini memerlukan hidrolisis ATP (bukan kecerunan RANGTP) untuk menubuhkan pembentukan semula arah RNPM, yang akan mencapai sitoplasma.
Kompleks liang nuklear dan kawalan ekspresi gen
Beberapa kajian menunjukkan bahawa komponen CPN dapat mempengaruhi peraturan ekspresi genetik melalui kawalan struktur kromatin dan aksesnya kepada faktor transkripsi.
Dalam eukariota evolusi yang lebih baru, heterochromatin lebih disukai terletak di pinggir nukleus. Wilayah ini terganggu oleh saluran euchromatin, yang dikekalkan oleh bakul nuklear CPN. Persatuan Keranjang Nuklear dengan Euchromatin berkaitan dengan transkripsi gen.
Sebagai contoh, pengaktifan transkripsi di peringkat CPN membayangkan interaksi komponen bakul nuklear dengan protein seperti faktor eksport acetyltransferase histon dan RNA.
Oleh itu, bakul nuklear adalah platform untuk pelbagai gen penyelenggaraan (Pengemasan) sangat ditranskripsikan dan gen sangat disebabkan oleh perubahan dalam keadaan persekitaran.
Kompleks liang nuklear dan virologi
Jangkitan sel eukaryot virus bergantung pada cpn. Dalam setiap kes jangkitan virus, kejayaannya bergantung kepada DNA, RNA atau RPN melalui CPN untuk mencapai matlamat utama, iaitu replikasi virus.
Boleh melayani anda: Kemasukan sitoplasma: Apa, Ciri, FungsiAPE Virus 40 (SV40) telah menjadi salah satu model yang paling banyak dikaji untuk menyiasat fungsi CPN dalam translocalization dalam nukleus. Ini kerana SV40 mempunyai genom kecil (5.000 pangkalan).
Telah ditunjukkan bahawa pengangkutan DNA virus memudahkan protein protein virus, yang melindungi virus sehingga nukleus mencapai.
Komponen
CPN tertanam di dalam sampul nuklear dan terdiri daripada kira -kira 500 dan 1000 nupts. Protein ini dianjurkan dalam subkompleks struktur atau modul, yang berinteraksi antara satu sama lain.
Modul pertama adalah komponen pusat, atau cincin, di dalam liang dalam bentuk jam pasir, yang dibatasi oleh cincin diameter 120 nm yang lain di kedua -dua belah, intranuklear dan sitoplasma. Modul kedua ialah cincin nukleus dan sitoplasma (diameter 120 nm setiap satu) yang terletak di sekitar komponen berbentuk sandwat.
Modul ketiga adalah lapan filamen yang diproyeksikan dari cincin 120 nm dalam nukleoplasma dan membentuk struktur berbentuk bakul. Modul keempat terdiri daripada filamen yang diproyeksikan ke sisi sitoplasma.
Kompleks berbentuk y, yang terdiri daripada enam protein Nups dan Seh 1 dan sec 13, adalah kompleks terbesar dan terbaik CNP. Kompleks ini adalah unit penting yang merupakan sebahagian daripada perancah CPN.
Walaupun persamaan yang rendah antara urutan NUPS, perancah CPN sangat dipelihara dalam semua eukariot.
Rujukan
- Beck, m., Sakit, e. 2016. Kompleks liang nuklear: Memahami fungsinya melalui wawasan struktur. Ulasan Alam, Biologi Sel Molekul, DOI: 10.1038/nrm.2016.147.
- Ibarra, a., Hetzer, m.W. 2015. Protein liang nuklear dan kawalan fungsi genom. Gen dan Pembangunan, 29, 337-349.
- KABACHINSKI, g., Schwartz, t.Atau. 2015. Kompleks liang nuklear - struktur dan fungsi sekilas. Jurnal Sains Sel, 128, 423-429.
- Knocknhauer, k.Dan., Schwartz, t.Atau. 2016. Kompleks liang nuklear sebagai pintu yang fleksibel dan dinamik. Sel, 164, 1162-1171.
- Ptak, c., Aitchison, J.D., Wozniak, r.W. 2014. Nuklear pelbagai fungsi oleh kompleks: platform untuk mengawal ekspresi gen. Pendapat semasa Biologi Sel, DOI: 10.1016/j.CEB.2014.02.001.
- Stawicki, s.P., Steffen, j. 2017. Republikasi: Kompleks liang nuklear - Kajian komprehensif struktur dan fungsi. Jurnal Antarabangsa Perubatan Akademik, 3, 51-59.
- Tran, e.J., Wente, s.R. 2006. Nuklear Dinamik oleh Kompleks: Kehidupan di tepi. Sel, 125, 1041-1053.