

Peringkat fosforilasi oksidatif, fungsi dan inhibitor
- 4407
- 609
- Ms. Edgar Carroll
The Fosforilasi oksidatif Ia adalah proses di mana molekul ATP disintesis dari ADP dan pYo (Fosfat bukan organik). Mekanisme ini dilakukan oleh bakteria dan sel eukariotik. Dalam sel eukariotik, fosforilasi dijalankan dalam matriks mitokondria sel -sel bukan fotosintesis.
Pengeluaran ATP diarahkan oleh pemindahan elektron dari koenzim NADH atau FADH2 ke o2. Proses ini mewakili pengeluaran tenaga terbesar di dalam sel dan diperolehi daripada kemerosotan karbohidrat dan lemak.
Sumber: Robot8a [cc by-sa 4.0 (https: // creativeCommons.Org/lesen/by-sa/4.0)] Tenaga yang disimpan dalam gradien beban dan pH, juga dikenali sebagai daya motor protonik, membolehkan proses ini dijalankan. Kecerunan proton yang dihasilkan, menyebabkan bahagian luaran membran mempunyai beban positif kerana kepekatan proton (h+) dan matriks mitokondria negatif.
[TOC]
Di mana fosforilasi oksidatif berlaku?
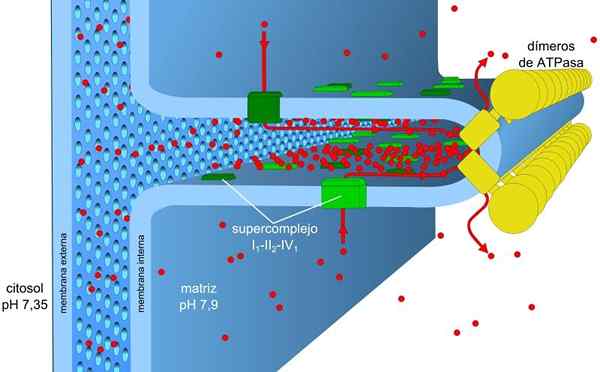
Proses pengangkutan elektron dan fosforilasi oksidatif dikaitkan dengan membran. Dalam prokariot mekanisme ini dijalankan melalui membran plasma. Dalam sel eukariotik mereka dikaitkan dengan membran mitokondria.
Bilangan mitokondria yang terdapat di dalam sel berbeza mengikut jenis sel. Sebagai contoh, dalam erythrocytes mamalia kekurangan organel ini, manakala jenis sel lain, seperti sel otot, boleh berjuta -juta mereka.
Membran mitokondria terdiri daripada membran luaran yang mudah, membran dalaman yang agak kompleks, dan di tengah -tengah mereka ruang intermembranal, di mana banyak enzim bergantung kepada ATP terletak.
Membran luar mengandungi protein yang dipanggil porina yang membentuk saluran untuk penyebaran mudah molekul kecil. Membran ini bertanggungjawab untuk mengekalkan struktur dan bentuk mitokondria.
Membran dalaman mempunyai ketumpatan yang lebih besar dan kaya dengan protein. Ia juga kalis air untuk molekul dan ion jadi, untuk menyeberang, mereka memerlukan protein intermembranal yang mengangkut mereka.
Di dalam matriks, lipatan membran dalaman memanjangkan, membentuk rabung yang membolehkannya mempunyai kawasan yang besar dalam jumlah kecil.
Tenaga Tenaga Sel
Mitokondria dianggap sebagai Pusat Tenaga Selular. Di dalamnya adalah enzim yang terlibat dalam proses kitaran asid cytric, pengoksidaan asid lemak dan enzim redoks dan protein pengangkutan elektron dan fosforilasi PhD.
Kecerunan kepekatan proton (kecerunan pH) dan kecerunan beban atau potensi elektrik dalam membran dalaman mitokondria, adalah penyebab daya motor protonik. Kebolehtelapan kecil membran dalaman untuk ion (selain daripada h+) membolehkan mitokondria mempunyai kecerunan voltan yang stabil.
Pengangkutan elektronik, proton mengepam dan ATP memperoleh serentak dalam mitokondria, terima kasih kepada daya motor protonik. Kecerunan pH mengekalkan keadaan berasid dalam matriks intermembrane dan mitokondria dengan keadaan alkali.
Untuk setiap dua elektron dipindahkan ke O2 Kira -kira 10 proton dipam oleh membran, mewujudkan kecerunan elektrokimia. Tenaga yang terpisah dalam proses ini dihasilkan secara beransur -ansur dengan melewati elektron melalui rantai penghantar.
Peringkat
Tenaga yang dikeluarkan semasa tindak balas pengurangan oksida NADH dan FADH2 Ia cukup tinggi (kira -kira 53 kcal/mol untuk setiap pasangan elektron), jadi untuk digunakan dalam pembuatan molekul ATP, ia mesti secara beransur -ansur dihasilkan dengan laluan elektron melalui pengangkut.
Ini dianjurkan di empat kompleks yang terletak di membran mitokondria dalaman. Gandingan tindak balas ini terhadap sintesis ATP dijalankan di kompleks kelima.
Boleh melayani anda: l limfosit b: ciri, struktur, fungsi, jenisRantaian Pengangkutan Elektron
NADH memindahkan beberapa elektron yang memasuki kompleks I rantaian pengangkutan elektron. Elektron dipindahkan ke mononukleotida Flavina, dan kemudian ke ubiquinone (Coenzyme Q) melalui penghantar besi-gula. Proses ini mengeluarkan sejumlah besar tenaga (16.6 kcal/mol).
Ubiquinona mengangkut elektron melalui membran ke kompleks III. Di kompleks ini elektron melalui cytochromes b dan c1 Terima kasih kepada pengangkut besi gula.
Dari kompleks III, elektron lulus ke kompleks IV (cytochrome c oxidase), dipindahkan satu oleh cytochrome C (protein membran periferal). Di kompleks IV, elektron melewati sepasang ion tembaga (CUke2+), kemudian ke cytochrome cke, Kemudian ke sepasang ion tembaga lain (CUb2+) dan dari ini ke cytochrome ke3.
Akhirnya, elektron dipindahkan ke O2 yang merupakan penerima terakhir dan membentuk molekul air (h2O) Bagi setiap pasangan elektron yang diterima. Laluan elektron dari kompleks IV ke O2 Ia juga menjana banyak tenaga percuma (25.8 kcal/mol).
Succinato Coq reductase
Kompleks II (Coq reductase succinate) menerima sepasang elektron dari kitaran asid sitrik, kerana pengoksidaan molekul succinate untuk fumarate. Elektron ini dipindahkan ke fad, kemudian melalui kumpulan besi-azufre, ke ubiquinona. Dari koenzim ini mereka pergi ke kompleks III dan ikuti laluan yang diterangkan sebelumnya.
Tenaga yang dikeluarkan dalam tindak balas pemindahan elektron ke FAD tidak mencukupi untuk meningkatkan proton melalui membran, sehingga dalam langkah ini rantai tidak ada daya motor protonik, dan akibatnya FADH menghasilkan kurang h+ daripada nadh.
Gandingan tenaga atau transduksi
Tenaga yang dihasilkan dalam proses pengangkutan elektron yang digambarkan sebelum ini, harus digunakan untuk pengeluaran ATP, reaksi yang dikatal oleh sintesis ATP atau enzim kompleks v. Pemuliharaan tenaga sedemikian dikenali sebagai gandingan tenaga, dan mekanisme sukar untuk dicirikan.
Beberapa hipotesis telah diterangkan untuk menggambarkan transduksi tenaga ini. Yang terbaik diterima ialah hipotesis gandingan chemosmotik, yang diterangkan di bawah.
Gandingan chemosmotic
Mekanisme ini mencadangkan bahawa tenaga yang digunakan untuk sintesis ATP berasal dari kecerunan protonik dalam membran sel. Proses ini campur tangan dalam mitokondria, kloroplas dan bakteria dan dikaitkan dengan pengangkutan elektron.
Kompleks pengangkutan elektronik I dan IV bertindak sebagai pam proton. Ini mengalami perubahan konformasi yang membolehkan mereka mengepam proton ke ruang intermembranal. Dalam kompleks IV untuk setiap pasangan elektron, dua proton dipam di luar membran dan dua lagi kekal dalam matriks yang membentuk h2Sama ada.
Ubiquinona di kompleks III menerima proton kompleks I dan II dan membebaskan mereka di luar membran. Kompleks I dan III masing -masing membenarkan laluan empat proton untuk setiap pasangan elektron yang diangkut.
Matriks mitokondria mempunyai kepekatan proton yang rendah dan potensi elektrik negatif manakala ruang intermembranal membentangkan keadaan songsang. Aliran proton melalui membran ini membayangkan kecerunan elektrokimia yang menyimpan tenaga yang diperlukan (± 5 kcal/mol oleh proton) untuk sintesis ATP.
Sintesis ATP
Enzim sintetik ATP adalah kompleks kelima yang terlibat dalam fosforilasi oksidatif. Bertanggungjawab untuk memanfaatkan tenaga kecerunan elektrokimia untuk membentuk ATP.
Protein transmembra ini terdiri daripada dua komponen: f0 dan f1. Komponen f0 Membolehkan kembalinya proton ke matriks mitokondria berfungsi sebagai saluran dan f F1 memangkinkan sintesis ATP melalui ADP dan PYo, Menggunakan tenaga pulangan tersebut.
Boleh melayani anda: protein penghantar membran: fungsi dan jenisProses sintesis ATP memerlukan perubahan struktur dalam f1 dan perhimpunan komponen f0 dan f1. Translocation of proton melalui f0 Ia menyebabkan perubahan konformasi dalam tiga subuniti f f1, membenarkan ini bertindak sebagai enjin putaran, mengarahkan pembentukan ATP.
Subunit yang bertanggungjawab untuk kesatuan ADP dengan pYo pergi dari keadaan lemah (l) ke aktif (t). Apabila ATP dibentuk, subunit kedua berlalu ke negeri terbuka (O) yang membolehkan pembebasan molekul ini. Selepas ATP dibebaskan, subunit ini berlalu dari negeri terbuka ke keadaan tidak aktif (l).
Molekul ADP dan PYo Mereka menyertai subunit yang telah pergi dari negeri atau negeri l.
Produk
Rantai penghantar elektron dan fosforilasi menghasilkan molekul ATP. Pengoksidaan NADH menghasilkan kira -kira 52.12 kcal/mol (218 kJ/mol) tenaga bebas.
Reaksi global untuk pengoksidaan NADH adalah:
Nadh+1/2 atau2 +H+ ↔ h2O+nad+
Pemindahan elektron dari NADH dan FADH2 Ia berlaku melalui beberapa kompleks, yang membolehkan perubahan tenaga bebas ΔG ° dibahagikan kepada "pakej" yang lebih kecil, yang ditambah pula dengan sintesis ATP.
Pengoksidaan molekul NADH menghasilkan sintesis tiga molekul ATP. Walaupun pengoksidaan molekul FADH2 Ia dilampirkan pada sintesis dua ATP.
Koenzim ini berasal dari proses glikolisis dan kitaran asid sitrik. Bagi setiap molekul glukosa yang terdegradasi, molekul 36 atau 38 ATP berakhir, bergantung kepada lokasi sel. Di otak dan otot rangka 36 ATP dihasilkan manakala 38 ATP dihasilkan dalam tisu otot.
Fungsi
Semua organisma, uniselular dan multiselular, memerlukan tenaga minimum dalam sel -sel mereka untuk menjalankan proses di dalamnya, dan seterusnya mengekalkan fungsi penting dalam badan yang lengkap.
Proses metabolik memerlukan tenaga untuk dijalankan. Kebanyakan tenaga yang boleh digunakan diperolehi oleh kemerosotan karbohidrat dan lemak. Tenaga ini berasal dari proses fosforilasi oksidatif.
Kawalan fosforilasi oksidatif
Kadar penggunaan ATP dalam sel, mengawal sintesis yang sama, dan seterusnya, disebabkan oleh gandingan fosforilasi oksidatif dengan rantai pengangkutan elektron, ia juga secara umumnya mengawal kadar pengangkutan elektronik.
Fosforilasi oksidatif mempunyai kawalan ketat yang memastikan ATP tidak dihasilkan lebih cepat daripada yang dimakan. Terdapat langkah -langkah tertentu dalam proses pengangkutan elektron dan fosforilasi yang digabungkan yang mengawal kadar pengeluaran tenaga.
Kawalan yang diselaraskan terhadap pengeluaran ATP
Laluan pengeluaran tenaga utama (sel ATP) adalah glikolisis, kitaran asid sitrik dan fosforilasi oksidatif. Kawalan yang diselaraskan dari tiga proses ini mengawal sintesis ATP.
Kawalan fosforilasi oleh nisbah tindakan massa ATP bergantung kepada sumbangan elektron yang tepat dalam rantai penghantar. Ini seterusnya bergantung pada hubungan [nadh]/[nad+] yang dipelihara ditinggikan oleh tindakan glikolisis dan kitaran asid sitrik.
Kawalan yang diselaraskan ini dijalankan dengan mengawal selia titik kawalan glikolisis (sitrat menghalang PFK) dan kitaran asid cytric (dehidrogenase piruvat, memotong sitrat, gatal dehidrogenase dan α-zetoglutarate dehydrogenase).
Kawalan oleh penerima
Kompleks IV (cytochrome c oxidase) adalah enzim yang dikawal oleh salah satu substratnya, iaitu kegiatannya dikawal oleh sitokrom yang dikurangkan C (c (c2+), yang seterusnya seimbang dengan hubungan kepekatan antara [nadh]/[nad+] dan nisbah tindakan massa [ATP]/[ADP] + [pYo].
Ia boleh melayani anda: nada: ciri dan fungsiSemakin tinggi [nadh]/[nad+] dan menurunkan [ATP]/[ADP]+[PYo], lebih banyak kepekatan akan menjadi cytochrome [c2+] dan lebih besar akan menjadi aktiviti kompleks IV. Ini ditafsirkan, sebagai contoh, jika kita membandingkan organisma dengan aktiviti berehat yang berbeza dan aktiviti yang tinggi.
Dalam individu dengan aktiviti fizikal yang tinggi, penggunaan ATP dan oleh itu hidrolisisnya kepada ADP + PYo Ia akan menjadi sangat tinggi, menghasilkan perbezaan dalam nisbah tindakan massa yang menyebabkan peningkatan [c2+dan oleh itu peningkatan sintesis ATP. Dalam individu berehat keadaan terbalik berlaku.
Pada akhirnya, kelajuan fosforilasi oksidatif meningkat dengan kepekatan ADP dalam mitokondria. Kepekatan ini bergantung kepada translocator ADP-ATP yang bertanggungjawab terhadap pengangkutan nukleotida adenine dan pYo Dari sitosol ke matriks mitokondria.
Ejen decacoplants
Fosforilasi oksidatif dipengaruhi oleh agen kimia tertentu, yang membolehkan pengangkutan elektronik diteruskan tanpa fosforilasi ADP, mereputasi pengeluaran dan pemuliharaan tenaga.
Ejen -ejen ini merangsang kelajuan penggunaan oksigen mitokondria jika tiada ADP, juga menyebabkan peningkatan dalam hidrolisis ATP. Mereka bertindak dengan menghapuskan perantara atau melanggar keadaan tenaga dari rantai penghantar elektron.
2.4-dinitrofenol, asid lemah yang melewati membran mitokondria, bertanggungjawab untuk menghilangkan kecerunan proton, kerana mereka menyertai mereka di sisi berasid dan melepaskannya di sisi asas.
Kompaun ini digunakan sebagai "pil pelangsingan" kerana didapati bahawa ia menghasilkan peningkatan pernafasan, oleh itu, peningkatan kadar metabolik dan penurunan berat badan yang berkaitan. Walau bagaimanapun, ditunjukkan bahawa kesan negatifnya boleh menyebabkan kematian.
Pelepasan kecerunan protonik menghasilkan haba. Sel -sel tisu adiposa coklat menggunakan decoupling, dikawal secara hormon, untuk menghasilkan haba. Mamalia Hibernate dan bayi yang baru lahir yang kekurangan rambut terdiri daripada tisu ini yang berfungsi sebagai selimut haba.
Perencat
Sebatian atau ejen menghalang kedua -dua penggunaan atau2 (pengangkutan elektronik) seperti fosforilasi oksidatif yang berkaitan. Ejen -ejen ini mengelakkan pembentukan ATP melalui penggunaan tenaga yang dihasilkan dalam pengangkutan elektronik. Oleh itu, rantai pengangkutan berhenti kerana penggunaan tenaga tersebut tidak diatur.
Antibiotik Oligomicin berfungsi sebagai perencat fosforilasi dalam banyak bakteria, mencegah rangsangan ADP dari sintesis ATP.
Terdapat juga ejen ionofor, yang membentuk kompleks larut lemak dengan kation seperti k+ dan na+, dan melalui membran mitokondria dengan kation ini. Mitokondria kemudian menggunakan tenaga yang dihasilkan dalam pengangkutan elektronik untuk mengepam kation dan bukannya mensintesis ATP.
Rujukan
- Alberts, b., Bray, d., Hopkin, k., Johnson, a., Lewis, J., Raff, m., Roberts, k. & Walter, p. (2004). Biologi sel penting. New York: Sains Garland.
- Cooper, g. M., Hausman, r. Dan. & Wright, n. (2010). Sel. (ms. 397-402). Marbán.
- Devlin, t. M. (1992). Buku Teks Biokimia: Dengan Korelasi Klinikal. John Wiley & Sons, Inc.
- Garrett, r. H., & Grisham, c. M. (2008). Biokimia. Thomson Brooks/Cole.
- Lodish, h., Darnell, J. Dan., Berk, a., Kaiser, c. Ke., Krieger, m., Scott, m. P., & Matsudaira, p. (2008). Biologi sel mollecular. Macmillan.
- Nelson, d. L., & Cox, m. M. (2006). Prinsip Biokimia Lehninger Edisi ke -4. Ed Omega. Barcelona.
- Voet, d., & Voet, j. G. (2006). Biokimia. Ed. Pan -American Medical.
- « Ciri -ciri Euglenophyta, Pembiakan, Pemakanan, Klasifikasi
- Latihan, fungsi dan jenis/kumpulan glukosida »